蓄“刺”待发 一切从刺突蛋白开始。每个新冠病毒颗粒(virion)的表面有24-40个任意排列的刺突蛋白,这些刺突蛋白是病毒与人细胞融合的关键[2]。与流感病毒等其他病毒表面的刚性融合蛋白相比,新冠病毒的刺突蛋白非常灵活,能在三个点像铰链一样活动,德国马克斯·普朗克生物物理学研究所生物化学家Martin Beck和同事在2020年8月发表的一项研究中指出[3]。 如此一来,刺突蛋白就能任意掉头、摇摆、转动,这有利于它对细胞表面进行扫视,或是用多个刺突蛋白与一个人体细胞相结合。目前其他冠状病毒尚无类似实验数据,但由于刺突蛋白序列在演化上高度保守,可以推测所有冠状病毒都具有这一特点,Beck说。 研究人员在疫情早期就证实了新冠病毒刺突蛋白的RBD能与ACE2受体结合。ACE2受体是一种常见的蛋白,广泛分布于人类喉部和肺部的大部分细胞表面。这个受体还是导致严重急性呼吸系统综合征的SARS病毒(SARS-CoV)的入胞点。但是,新冠病毒与ACE2的亲和力更好,估计是SARS病毒的2-4倍[4],这是因为RBD的多种变化能稳定其与病毒结合的热点位置[5]。 危险的新冠病毒变异株容易在刺突蛋白的S1亚基上携带突变,S1亚基包含RBD,并负责病毒与ACE2受体的结合(刺突蛋白的S2亚基则能促进病毒外膜与宿主细胞膜的融合。) 以Alpha变异株为例,它在刺突蛋白序列上有10个突变,导致RBD更有可能保持“向上”的状态[6]。“这能让病毒更易进入细胞。”美国北卡罗来纳州杜克人类疫苗研究所的结构生物学家Priyamvada Acharya说。Acharya正在研究刺突蛋白的突变。 目前Delta变异株正在世界各地传播,它在S1亚基上有多个突变,包括RBD上的3个突变,这3个突变似乎不仅能提高RBD与ACE2的亲和力,还能提高其逃逸免疫系统的能力[7]。
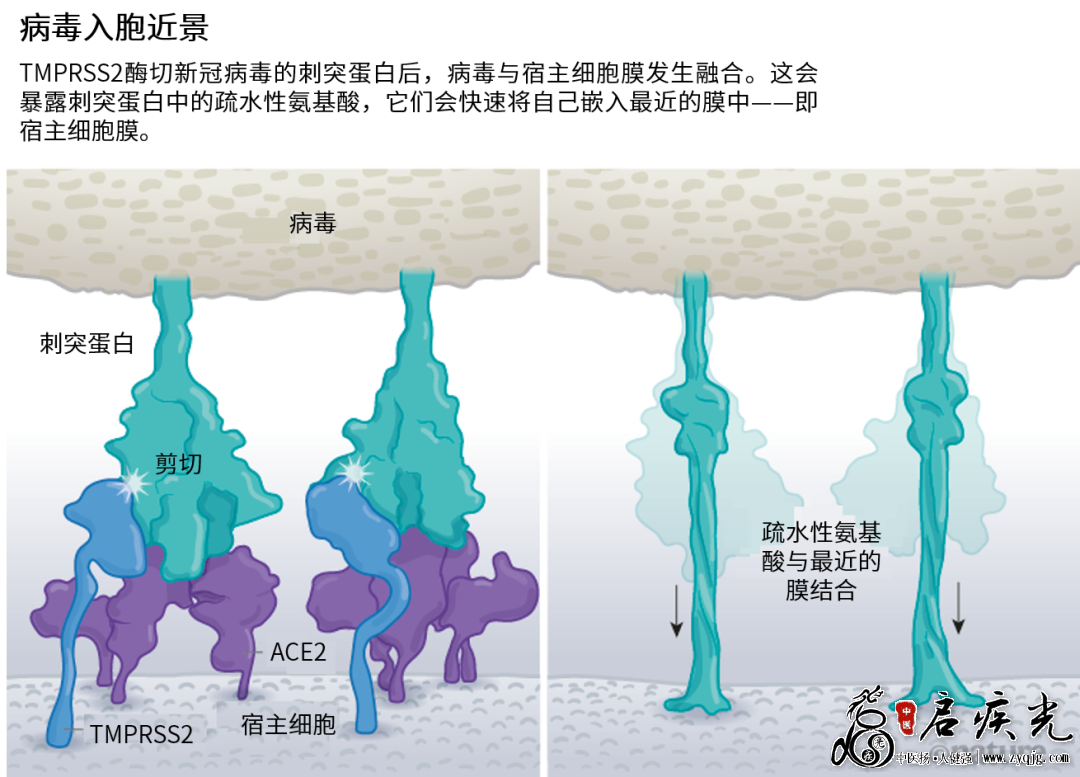
限定入口 病毒刺突蛋白与ACE2结合后,宿主细胞表面的其他蛋白就会启动病毒外膜与细胞膜融合的过程(见“病毒入胞近景”)。
来源:Janet Iwasa, Univ. Utah;图:Nik Spencer/Nature SARS病毒会利用两个宿主蛋白酶中的一个入胞:TMPRSS2(读作tempress two)或组织蛋白酶L。TMPRSS2的入胞途径更快,但SARS病毒经常通过核内体侵入,核内体是一种脂质包裹的囊泡,这个途径依赖组织蛋白酶L。不过,如果病毒颗粒从这一途径进入,就会被抗病毒蛋白逮个正着。 新冠病毒之所以不同于SARS病毒,就在于它能更快地利用TMPRSS2。TMPRSS2是呼吸道细胞表面大量存在的一种酶。TMPRSS2会先酶切刺突蛋白S2亚基上的一个位点[8]。这个剪切点会暴露一串疏水性氨基酸,暴露后的疏水性氨基酸会迅速嵌入最近的膜中——即宿主细胞膜。随后,展开的刺突蛋白会折叠起来,像拉链一样,迫使病毒外膜与细胞膜融合。 随后,新冠病毒将基因组直接注射到宿主细胞内。通过采取这种弹簧式的侵入方式,新冠病毒比SARS病毒的感染更快,而且不会被核内体逮住,Barclay和她在英国帝国理工学院的同事在4月发表的一项研究中描述道[9]。 新冠病毒能利用TMPRSS2实现快速入胞,解释了为何疟疾药物氯喹一开始在实验室研究中表现良好,但在治疗COVID-19的临床试验中无效[10]。原来氯喹利用的细胞完全依赖组织蛋白酶实现核内体侵入。“新冠病毒在人类气道中传播和复制时不会用到核内体,所以氯喹这种核内体干扰药物在真实人体中效果就不大了。”Barclay说。 这一发现还指出,蛋白酶抑制剂是一种很有希望的治疗选项,可以防止病毒利用TMPRSS2、组织蛋白酶L或其他蛋白酶进入宿主细胞。camostat mesylate是一种TMPRSS2抑制剂,日本已将其批准用于治疗胰腺炎。这种抑制剂能阻断病毒进入肺部细胞[8],但无法在初步临床试验中提高病人的治疗转归[11]。 “依我看,我们应当将这种蛋白酶抑制剂作为广谱抗病毒药物,防止新的疾病发展为大流行,将其遏制在萌芽阶段。”德国灵长类研究中心感染生物学部主任Stefan Pöhlmann说。Pöhlmann领导开展了ACE2结合和TMPRSS2途径的研究。
致命竞争 接下来的感染步骤就没那么清晰了。“进入细胞之后就有很多黑箱。不确定性和各种假说也更多了。”美国犹他大学的化学家Janet Iwasa说。Iwasa正在制作一个解释新冠病毒生命周期的带注释的动画。 就在新冠病毒将RNA基因组注射到宿主细胞后,细胞质核糖体会将两个病毒RNA片段翻译成氨基酸长链,这些氨基酸长链再被切割出16个蛋白质,包括许多参与RNA合成的蛋白质。随后会产生更多的RNA,这些RNA编码26个已知的病毒蛋白,包括用来制造新病毒颗粒的结构蛋白(比如刺突蛋白)和其他协助蛋白。这样,病毒就能大量产生其自身的信使RNA(mRNA)拷贝,但它还需要细胞机器来将这些mRNA翻译成蛋白质。 新冠病毒有很多将细胞机器占为己用的策略。病毒学家Noam Stern-Ginossar和她在以色列魏茨曼科学研究所的团队重点研究了新冠病毒抑制宿主mRNA翻译、促进自身mRNA翻译的三种机制。虽然这三种机制并非新冠病毒所独有,但这些作用的结合、速度和程度看来确实是独一无二的,Stern-Ginossar说。 第一,新冠病毒会清除竞争对手:病毒蛋白Nsp1是新冠病毒抵达时首先被翻译的蛋白之一,它会把宿主蛋白质招募起来,系统性地切割所有不带病毒标记的细胞mRNA。如果Stern-Ginossar的团队将同样的病毒标记放在宿主mRNA的末端,这个mRNA就不会被切割[12]。 第二,感染会让细胞内全部蛋白质翻译减少70%。Nsp1依然是捣蛋分子,这一次它能阻断核糖体的入胞渠道,让mRNA无法进入,两个研究团队分别得出了以上结论[13,14]。剩下不多的翻译能力全被用来翻译病毒RNA,Stern-Ginossar说。 第三,新冠病毒会关闭细胞的预警系统。这有很多方式,但Stern-Ginossar的团队发现了新冠病毒的一个明确机制:病毒让细胞mRNA无法离开细胞核,包括提醒免疫系统注意感染的蛋白质的指令。另一个团队证实了这个结果,再次将矛头指向Nsp1:这个蛋白似乎会阻塞离开细胞核的通道,一个不让逃走[15]。 由于基因转录本无法离开细胞核,因此受感染的细胞不会释放许多干扰素——干扰素是提醒免疫系统注意病毒的信号蛋白。新冠病毒关闭这一预警系统的速度尤其快:和其他呼吸道病毒相比,包括SARS病毒和呼吸道合胞病毒,新冠病毒感染后诱导的干扰素水平显著降低[16]。今年6月,研究人员报道了Alpha变异株的突变似乎能更有快地减少干扰素的产生[17]。 “新冠病毒明显是个动作很快的病毒,具有非常独特的能力,能从源头上扰乱免疫系统识别病毒和抵抗感染的能力。”Stern-Ginossar说。等到免疫系统真正发现有病毒时,病毒数量已经太多了,致使免疫应答蛋白有时会比平常更快地充斥在血液中,而这可能对人体有害。临床医生在疫情很早就发现,有些发展成重症的COVID-19患者除了受到病毒本身的袭击外,过度激活的免疫应答也造成了一定伤害。一些经证明有效的疗法专门抑制这种免疫应答。
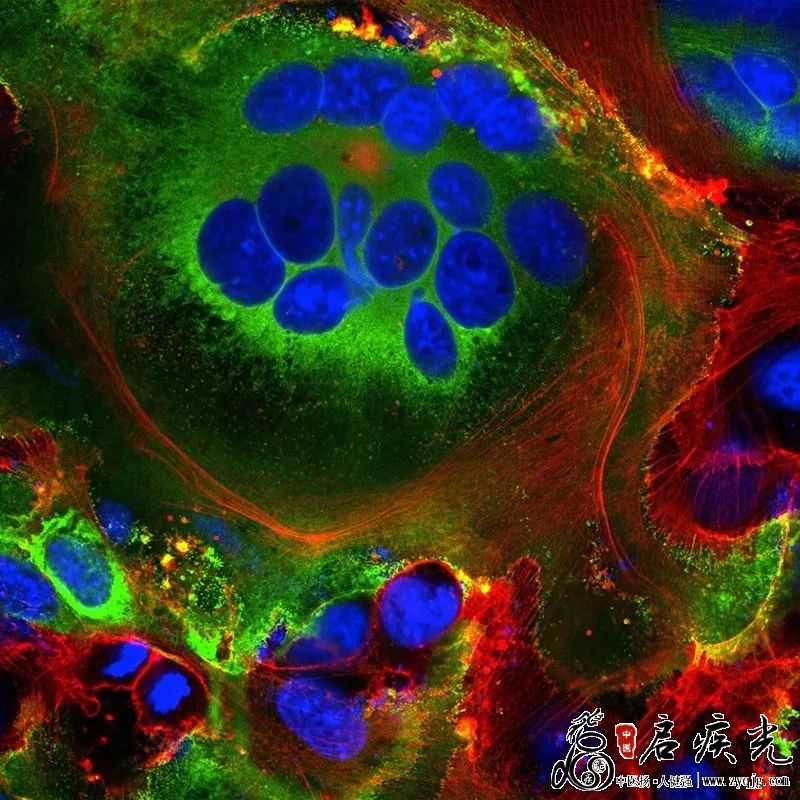
最强改造王 新冠病毒接管宿主细胞翻译后,它就开始喧宾夺主了,朝着有利自己的方向大肆改造细胞内部和表面。 首先,一些新制造的刺突蛋白会抵达宿主细胞表面,突破宿主细胞膜。它们还会在那里激活一条宿主钙离子通道,在细胞表面分泌一层脂肪膜——肌肉细胞等自然融合的细胞上也会发现这种膜。这时,受感染细胞与表达ACE2的相邻细胞融合,发展为最多包含20个细胞核的单个大呼吸道细胞。
表达新冠病毒刺突蛋白(绿色)的细胞内可见融合细胞结构(合胞体)。蓝色为细胞核,红色为细胞骨架。来源:Mauro Giacca
|